Биология в лицее
Site biology teachers lyceum № 2 Voronezh city, Russian Federation
2. Питательная — переносят на своей поверхности аминокислоты и доставляют их клеткам организма.
3. Буферная — поддержание с помощью гемоглобина рН крови.
4. Регуляторная — переносят на своей поверхности витамины, гормоны и различные ферменты.
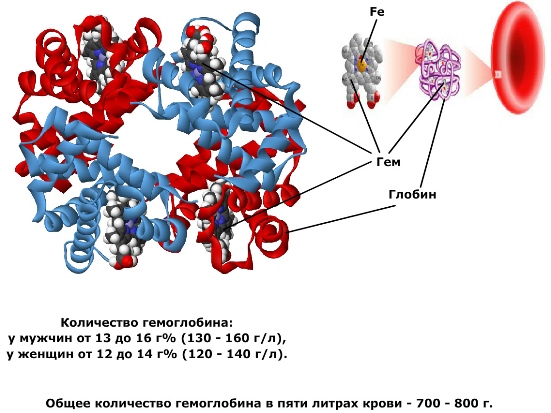
Гемоглобин — сложное химическое соединение, которое находится в эритроцитах и состоит из двух частей: белка глобина и четырёх молекул гема. Таким образом, гемоглобин — это сложный белок — хромопротеид. Гем имеет в своём составе атом железа, способный присоединять и отдавать молекулу кислорода. При этом валентность железа не изменяется (оно остаётся двухвалентным).
В капиллярах лёгких гемоглобин присоединяет кислород и становится оксигемоглобином, который придаёт артериальной (богатой кислородом) крови ярко-алый цвет. В капиллярах тканей гемоглобин отдаёт кислород и превращается в восстановленный гемоглобин, который придаёт венозной крови тёмно-вишнёвый цвет. При соединении гемоглобина с углекислым газом (CO2) образуется карбогемоглобин.
При разрушении эритроцитов гемоглобин выходит в плазму крови, которая окрашивается при этом в красный цвет и становится прозрачной — «лаковая кровь». Процесс разрушения эритроцитов и выхода гемоглобина носит название гемолиз. Он может произойти при уменьшении осмотического давления, что вначале приводит к набуханию, а затем разрушению эритроцитов, при воздействии химических веществ, разрушающих оболочку эритроцитов (эфир, хлороформ, алкоголь, желчные кислоты и т. д.), при сильных механических воздействиях на кровь, например при её транспортировке, при замораживании и размораживании крови, при её нагревании до 65 — 68 о С, при переливании несовместимой или недоброкачественной крови, при укусах ядовитых змей, скорпионов, при иммунных реакциях.
В скелетных мышцах и миокарде (сердечной мышце) находится мышечный гемоглобин — миоглобин. Он отличается от гемоглобина крови меньшей молекулярной массой глобина (белковой части). Назначение миоглобина — снабжение кислородом работающей мышцы в момент сокращения, когда кровоток в ней уменьшен или прекращается вовсе.
Эритроциты
| Эритроцит | ||
|---|---|---|
 |
Общий миелоидный прародитель → Проэритробласт → Мегалобласт → Полихроматический эритробласт → Нормоцит → Ретикулоцит → Эритроцит

Эритроци́ты (от греч. ἐρυθρός — красный и κύτος — вместилище, клетка), также известные под названием кра́сные кровяны́е тельца́, — клетки крови человека, позвоночных животных и некоторых беспозвоночных (сипункулид, у которых эритроциты плавают в полости целома [1] ).
Содержание
Функции
Эритроциты — высокоспециализированные клетки, функцией которых является перенос кислорода из лёгких к тканям тела и транспорт диоксида углерода (CO2) в обратном направлении. У позвоночных, кроме млекопитающих, эритроциты имеют ядро, у эритроцитов млекопитающих ядро отсутствует.
Наиболее специализированы эритроциты млекопитающих, лишённые в зрелом состоянии ядра и органелл и имеющие форму двояковогнутого диска, обуславливающую высокое отношение площади к объёму, что облегчает газообмен. Особенности цитоскелета и клеточной мембраны позволяют эритроцитам претерпевать значительные деформации и восстанавливать форму (эритроциты человека диаметром 8 мкм проходят через капилляры диаметром 2—3 мкм).
Транспорт кислорода обеспечивается гемоглобином (Hb), на долю которого приходится ≈98 % массы белков цитоплазмы эритроцитов (в отсутствии других структурных компонентов). Гемоглобин является тетрамером, в котором каждая белковая цепь несёт гем — комплекс протопорфирина IX с ионом двухвалентного железа, кислород обратимо кординируется с ионом Fe 2+ гемоглобина, образуя оксигемоглобин HbO2:
Hb + O2  HbO2
HbO2
Особенностью связывания кислорода гемоглобином является его аллостерическое регулирование — стабильность оксигемоглобина падает в присутствии 2,3-дифосфоглицериновой кислоты — промежуточного продукта гликолиза и, в меньшей степени, углекислого газа, что способствует высвобождению кислорода в тканях, в нём нуждающихся.
Транспорт углекислого газа эритроцитами происходит с участием карбоангидразы, содержащейся в их цитоплазме. Этот фермент катализирует обратимое образование бикарбоната из воды и углекислого газа, диффундирующего в эритроциты:
H2O + CO2 H + + HCO3 –
В результате в цитоплазме накапливаются ионы водорода, однако снижение pH при этом незначительно из-за высокой буферной ёмкости гемоглобина. Вследствие накопления в цитоплазме ионов бикарбоната возникает градиент концентрации, однако ионы бикарбоната могут покидать клетку только при условии сохранения равновесного распределения зарядов между внутренней и внешней средой, разделённых цитоплазматической мембраной, то есть выход из эритроцита иона бикарбоната должен сопровождаться либо выходом катиона, либо входом аниона. Мембрана эритроцита практически непроницаема для катионов, но содержит хлоридные ионые каналы, в результате выход бикарбоната из эритроцита сопровождается входом в него хлорида (хлоридный сдвиг).
Формирование эритроцитов
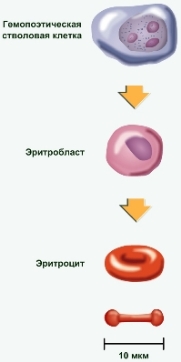
Формирование эритроцитов (эритропоэз) происходит в костном мозге черепа, рёбер и позвоночника, а у детей — ещё и в костном мозге в окончаниях длинных костей рук и ног. Продолжительность жизни — 3—4 месяца, разрушение (гемолиз) происходит в печени и селезёнке. Прежде чем выйти в кровь, эритроциты последовательно проходят несколько стадий пролиферации и дифференцировки в составе эритрона — красного ростка кроветворения.
Полипотентная стволовая клетка крови (СКК) даёт клетку-предшественницу миелопоэза (КОЕ-ГЭММ), которая в случае эритропоэза даёт клетку-родоначальницу миелопоэза (КОЕ-ГЭ), которая уже даёт унипотентную клетку, чувствительную к эритропоэтину (БОЕ-Э).
Бурстобразующая единица эритроцитов (БОЕ-Э) даёт начало эритробласту, который через образование пронормобластов уже дают морфологически различимые клетки-потомки нормобласты (последовательно переходящие стадии):
Гемопоэз (в данном случае эритропоэз) исследуется по методу селезёночных колоний.
Большая клетка с ядром, не обладающая характерным красным цветом, — мегалобласт; затем она окрашивается в красный цвет — теперь это эритробласт. Нормоцит (нормобласт) уменьшается в размере в процессе развития. После утраты ядра нормоцит превращается в ретикулоцит.
У птиц, пресмыкающихся, земноводных и рыб ядро просто теряет активность, но сохраняет способность реактивации. Одновременно с исчезновением ядра по мере взросления эритроцита из его цитоплазмы исчезают рибосомы и другие компоненты, участвующие в синтезе белка. Ретикулоциты попадают в кровеносную систему и через несколько часов становятся полноценными эритроцитами.
Структура и состав
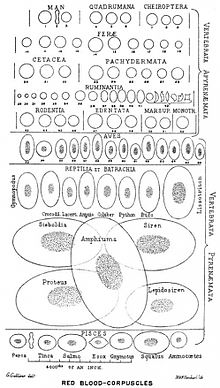

У большинства групп позвоночных эритроциты имеют ядро и другие органоиды.
У млекопитающих зрелые эритроциты лишены ядер, внутренних мембран и большинства органоидов. Ядра выбрасываются из клеток-предшественников в ходе эритропоэза. Обычно эритроциты млекопитающих имеют форму двояковогнутого диска и содержат в основном дыхательный пигмент гемоглобин. У некоторых животных (например, верблюда) эритроциты имеют овальную форму.
Содержимое эритроцита представлено главным образом дыхательным пигментом гемоглобином, обусловливающим красный цвет крови. Однако на ранних стадиях количество гемоглобина в них мало, и на стадии эритробластов цвет клетки синий; позже клетка становится серой и, лишь полностью созрев, приобретает красную окраску.

Важную роль в эритроците выполняет клеточная (плазматическая) мембрана, пропускающая газы (кислород, углекислый газ), ионы (Na, K) и воду. Плазмолемму пронизывают трансмембранные белки — гликофорины, которые, благодаря большому количеству остатков сиаловой кислоты, ответственны примерно за 60 % отрицательного заряда на поверхности эритроцитов.
В одном литре крови содержится эритроцитов:
Переливание крови
При переливании крови от донора к реципиенту возможна агглютинация (склеивание) и гемолиз (разрушение) эритроцитов. Чтобы этого не происходило, необходимо учитывать группы крови, открытые К. Ландштейнером и Я. Янским в 1900 году. Агглютинацию вызывают белки, находящиеся на поверхности эритроцита — антигены (агглютиногены) и находящиеся в плазме антитела (агглютинины). Существуют 4 группы крови, для каждой характерны различные антигены и антитела. Переливание обычно проводится лишь между обладателями одной группы крови.
| I — 0 | II — A | III — B | IV — AB |
|---|---|---|---|
| αβ | β | α | — |
Место в организме
Форма двояковогнутого диска обеспечивает прохождение эритроцитов через узкие просветы капилляров. В капиллярах они движутся со скоростью 2 сантиметра в минуту, что даёт им время передать кислород от гемоглобина к миоглобину. Миоглобин действует как посредник, принимая кислород у гемоглобина в крови и передавая его цитохромам в мышечных клетках.
Количество эритроцитов в крови в норме поддерживается на постоянном уровне (у человека в 1 мм³ крови 4,5—5 млн эритроцитов, у некоторых копытных 15,4 млн (лама) и 13 млн (коза) эритроцитов, у пресмыкающихся — от 500 тыс. до 1,65 млн, у хрящевых рыб — 90—130 тыс.) Общее число эритроцитов снижается при анемиях, повышается при полицитемии.
Продолжительность жизни эритроцита человека в среднем 125 суток (ежесекундно образуется около 2,5 млн эритроцитов и такое же их количество разрушается), у собак — 107 дней, у кроликов и кошек — 68.
Патология

При различных заболеваниях крови возможно изменение цвета эритроцитов, их размеров, количества, а также формы; они могут принимать, например, серповидную, овальную, сферическую или мишеневидную форму.
При изменении кислотно-щелочного баланса крови в сторону закисления (от 7,43 до 7,33) происходит склеивание эритроцитов в виде монетных столбиков, либо их агрегация.
Среднее содержание гемоглобина для мужчин 13,3—18 г% (или 4,0—5,0·10 12 единиц), для женщин 11,7—15,8 г% (или 3,9—4,7·10 12 единиц). Единица измерения уровня гемоглобина представляет собой процент содержания гемоглобина в 1 грамме эритроцитарной массы.
Что общего у эритроцитов млекопитающих
Используя содержание текста «Древние птицы Новой Зеландии» и знания школьного курса биологии, выполните задания и ответьте на вопрос.
1) Составьте наиболее вероятную пищевую цепь, которая сложилась на территории Новой Зеландии до появления на острове современного человека.
2) Вычислите примерное значение наибольшей массы добычи, которую могла переносить самка орла Хааста. Запишите арифметическое выражение и численный ответ (в килограммах).
3) Какой критерий использовали учёные при установлении родства орла Хааста с другими пернатыми хищниками?
ДРЕВНИЕ ПТИЦЫ НОВОЙ ЗЕЛАНДИИ
Новая Зеландия отделилась от Гондваны раньше Австралии, ещё в меловом периоде, и её уникальная фауна самая древняя в мире. На этом горном архипелаге, покрытом лесами, с вулканами и гейзерами, до появления человека совсем не было млекопитающих, не считая двух видов летучих мышей. Поэтому экологическую нишу травоядных копытных занимали мирные нелетающие птицы моа, родственники киви, напоминающие страусов с мощными ногами. Существовало не менее 20 видов моа, и только некоторые из них дожили до XIX века.
Мелкие моа были размером с индюка, а рост некоторых крупных достигал 3,5 м при массе 300–400 кг! Моа быстро не бегали: до появления человека бегать им было не от кого. Наземных хищников не имелось вовсе, только пернатые, а вершину пищевой пирамиды занимал орёл Хааста.
По ископаемым останкам скелетов учёные вычислили размеры и примерный вес этих птиц. Оказалось, что это самый крупный и тяжёлый из современных орлов, больше беркута и белохвоста, масса которых не превышает 7 кг. Размах крыльев орла Хааста достигал 2,1–2,4 м; масса самцов — 10 кг, а самок — 14,5 кг! Изучив пропорции его тела, учёные решили, что орёл Хааста совершенно непохож на парящих орлов — обитателей открытых просторов. У орла Хааста широкие и относительно недлинные крылья, как у лесных хищников, например у гарпий. Добыча же ему нужна была достаточно крупная, и среди кандидатов на роль жертвы учёные называют нелетающих пастушков, а также не очень больших моа, которых тяжёлый орёл, возможно, сбивал с ног, почти падая на них из крон деревьев, а потом убивал своими огромными когтями. Поскольку пернатые хищники способны поднять в воздух добычу, лишь на четверть превышающую их вес, вряд ли орёл питался 200-килограммовыми моа, однако вполне мог подкормиться их трупами и птенцами.
Совсем недавно учёные сравнили ДНК митохондрий 16 современных видов орлов с ДНК орла Хааста из ископаемых костей, возраст которых — 2 тыс. лет. По результатам этого теста ближайшим родственником нашего гиганта оказался орёл-карлик и другие мелкие лесные ястребиные орлы из того же рода, а отделение этого вида от общего предка произошло не так давно — 0,7–1,8 млн лет назад.
Биохимия крови
| Сайт: | Образовательный портал МБФ (ВолгГМУ) |
| Курс: | Нарушения метаболизма. Биохимия специализированных тканей. (Для клин.ордов КЛД) |
| Книга: | Биохимия крови |
Оглавление
1. Общие сведения
2. свойства крови
3. Состав крови. Гематокрит

3.1. Плазма крови
Пла́зма кро́ви (от греч. πλάσμα — нечто сформированное, образованное) — жидкая часть крови, в которой взвешены форменные элементы — вторая часть крови. Процентное содержание плазмы в крови составляет 52—61 %. Макроскопически представляет собой однородную несколько мутную (иногда почти прозрачную) желтоватую жидкость, собирающуюся в верхней части сосуда с кровью после осаждения форменных элементов. Гистологически плазма является межклеточным веществом жидкой ткани крови.
Центрифуги-сепараторы выделяют из крови плазму. Плазма крови состоит из воды, в которой растворены вещества — белки (7—8 % от массы плазмы) и другие органические и минеральные соединения. Основными белками плазмы являются альбумины — 55—65 %, α1-глобулины — 2—4 %, α2-глобулины 6—12 %, β-глобулины8 — 12 %, γ-глобулины — 2-4 % и фибриноген — 0,2—0,4 %. В плазме крови растворены также питательные вещества (в частности глюкоза и липиды), гормоны, витамины, ферменты и промежуточные и конечные продукты обмена веществ, а также неорганические вещества.
В среднем 1 литр плазмы человека содержит 900—950 г воды, 65—85 г белка и 20 г низкомолекулярных соединений. Плотность плазмы составляет от 1,025 до 1,029, pH — 7,36—7,44.
Существует обширная практика собирания донорской плазмы крови. Плазма отделяется от эритроцитов центрифугированием с помощью специального аппарата, после чего эритроциты возвращаются донору. Этот процесс называется плазмаферезом.
Плазма с высокой концентрацией тромбоцитов (богатая тромбоцитами плазма) находит все большее применение в медицине в качестве стимулятора заживления и регенерации тканей организма. В настоящее время на её основе разработана многофункциональная медицинская методика, используемая в стоматологии и косметологии.
3.2. Форменные элементы
У взрослого человека форменные элементы крови составляют около 40—50 %, а плазма — 50—60 %. Форменные элементы крови представлены эритроцитами, тромбоцитами и лейкоцитами:
Кровь относится к быстро обновляющимся тканям. Физиологическая регенерация форменных элементов крови осуществляется за счёт разрушения старых клеток и образования новых органами кроветворения. Главным из них у человека и других млекопитающих является костный мозг. У человека красный, или кроветворный, костный мозг расположен в основном в тазовых костях и в длинных трубчатых костях. Основным фильтром крови является селезёнка (красная пульпа), осуществляющая в том числе и иммунологический её контроль (белая пульпа).
4. Биохимия эритроцита
4.1. Транспорт кислорода кровью
4.2. Созревание эритроцита
4.3. Структурно-метаболические особенности эритроцита
Особенности структурной организации мембраны эритроцитов
Эритроцит окружен плазматической мембраной, структура которой хорошо изучена, идентична таковой в других клетках. Цитоплазматическая мембрана эритроцитов включает бислой фосфолипидов, в то время как белки или «плавают» на поверхности мембран, или пронизывают липиды, обеспечивая прочность и вязкость мембран. Площадь мембраны одного эритроцита составляет около 140 мкм2.
На долю белков приходится примерно 49 %, липидов – 44 %, углеводов –7 %. Углеводы химически связаны либо с белками, либо с липидами и образуют соответственно гликопротеиды и гликолипиды.
Важнейшими компонентами мембраны эритроцитов являются липиды, включающие до 48 % холестерина, 17-28 % – фосфотидилхолина, 13-25 % – сфингомиелина и ряд других фосфолипидов.
Фосфотидилхолин мембраны эритроцитов несет нейтральный заряд, практически не вступает в реакции взаимодействия с положительно заряженными каналами Са2+,, обеспечивая тем самым атромбогенность эритроцитов. Благодаря таким свойствам, как текучесть, пластичность, эритроциты способны проходить через капилляры диаметром
Белки мембраны эритроцита делят на периферические и интегральные. К периферическим белкам относят спектрин, анкирин, белок 4.1., белок р55, адуцин и др. В группу интегральных белков входит фракция 3, а также гликофорины А, В, С, О, Е. Анкирин образует соединение с р-спектрином. В составе эритроцитов обнаружено около 340 мембранных и 250 растворимых белков.
Пластичность эритроцитов связана с фосфорилированием мембранных белков, особенно белков полосы 4.1.
Белок фракции 4.2. – паллидин обеспечивает связывание спектрин-актин-анкиринового комплекса с фракцией 3, относится к группе трансглутаминазных протеинов.
К числу сократительных белков мембраны эритроцитов относятся р-актин, тропомодулин, строматин и тропомиозин.
Гликофорины – интегральные белки мембраны эритроцитов, определяющие отрицательный заряд, способствующий отталкиванию эритроцитов друг от друг и от эндотелия сосуда.
Протеин 3 – основной белок актинов, регулирующий дефосфорилируемость эритроцита.
Как указывалось выше, мембрана эритроцита представляет собой сложный комплекс, включающий определенным образом организованные липиды, белки и углеводы, которые формируют наружный, средний и внутренний слои эритроцитарной мембраны.
Касаясь пространственного расположения различных химических компонентов эритроцитарной мембраны, следует отметить, что наружный слой образован гликопротеидами с разветвленными комплексами олигосахаридов, которые являются концевыми отделами групповых антигенов крови. Липидным компонентом наружного слоя являются фосфатидилхолин, сфингомиелин и неэстерифицированный холестерин. Липиды наружного слоя мембраны эритроцита играют важную роль в обеспечении постоянства структуры мембраны, избирательности ее проницаемости для различных субстратов и ионов. Вместе с фосфолипидами холестерин регулирует активность мембранно-связанных ферментов путем изменения вязкости мембраны, а также участвует в модификации вторичной структуры ферментов. Молярное отношение холестерин / фосфолипиды в мембранах клеток у человека и многих млекопитающих равно 0,9. Изменение этого соотношения в сторону увеличения наблюдается в пожилом возрасте, а также при некоторых заболеваниях, связанных с нарушением холестеринового обмена.
Снижение текучести мембраны эритроцита и изменение ее свойств отмечается также и при увеличении содержания сфингомиелина,
Средний бислой мембраны эритроцита представлен гидрофобными «хвостами» полярных липидов. Липидный бислой обладает выраженной текучестью, которая обеспечивается определенным соотношением между насыщенными и ненасыщенными жирными кислотами гидрофобной части бислоя. Интегральные белки, к которым относятся ферменты, рецепторы, транспортные белки, обладают активностью только в том случае, если находятся в гидрофобной части бислоя, где они приобретают необходимую для активности пространственную конфигурацию. Поэтому любые изменения в составе липидов эритроцитарной мембраны сопровождаются изменением ее текучести и нарушением работы интегральных белков.
Внутренний слой мембраны эритроцита, обращенный к цитоплазме, состоит из белков спектрина и актина. Спектрин является специфическим белком эритроцитов, его гибкие вытянутые молекулы, связываясь с микрофиламентами актина и липидами внутренней поверхности мембраны, формируют своеобразный скелет эритроцита. Небольшой процент липидов во внутреннем слое мембраны эритроцита представлен фосфатидилэтаноламином и фосфатидилсерином. От наличия спектрина зависит подвижность белков, удерживающих двойной бисой липидов.
Одним из важных гликопротеинов является гликофорин, содержащийся как на внешней, так и на внутренней поверхностях мембран эритроцитов. Гликофорин в своем составе содержит большое количество сиаловой кислоты и обладает значительным отрицательным зарядом. В мембране он располагается неравномерно, образует выступающие из мембраны участки, которые являются носителями иммунологических детерминант.
Строение и состояние эритроцитарной мембраны, низкая вязкость нормального гемоглобина обеспечивают значительные пластические свойства эритроцитам, благодаря которым эритроцит легко проходит по капиллярам, имеющим вдвое меньший диаметр, чем сама клетка, и может принимать самые разнообразные формы. Другим периферическим мембранным белком эритроцитов является анкирин, образующий соединение с молекулой Р-спектрина.
Функции эритроцитарной мембраны
Мембрана эритроцитов обеспечивает регуляцию электролитного баланса клетки за счет активного энергозависимого транспорта электролитов или пассивной диффузии соединений по осмотическому градиенту.
В мембране эритроцитов имеются ионно-проницаемые каналы для катионов Na+, K+, для O2, CO2, Cl– HCO3–.
Транспорт электролитов через эритроцитарную мембрану и поддержание его мембранного потенциала обеспечивается энергозависимыми Na+, K+, Ca2+ – АТФ-азными системами.
Мембрана эритроцитов хорошо проницаема для воды при участии так называемых белковых и липидных путей, а также анионов, газообразных соединений и плохо проницаема для одновалентных катионов калия и натрия.
Белковый путь трансмембранного переноса воды обеспечивается при участии пронизывающего мембрану эритроцитов белка «полосы 3», а также гликофорина.
Молекулярная природа липидного пути переноса воды через эритроцитарную мембрану практически не изучена. Прохождение молекул небольших гидрофильных неэлектролитов через эритроцитарную мембрану осуществляется также, как и перенос воды, за счет белкового и липидного путей. Перенос мочевины и глицерина через мембрану эритроцита обеспечивается за счет ферментативных реакций.
Перенос органических анионов через эритроцитарную мембрану обеспечивается, как и транспорт неорганических анионов, при участии белка «полосы 3».
Эритроцитарная мембрана обеспечивает активный транспорт глюкозы, кинетика которого обеспечивается зависимостью Михаэлиса-Ментен. Важная роль в транспорте глюкозы через эритроцитарную мембрану отводится полипептиду полосы 4,5 (белки с ММ 55 кД – возможные продукты распада полипептида полосы 3). Высказывается предположение о наличии специфического липидного окружения у белков – переносчиков сахаров в эритроцитарной мембране.
Неравномерное распределение моновалентных катионов в системе эритроцит – плазма крови поддерживается при участии энергозависимой Na+-помпы, осуществляющей трансмембранный обмен ионов Na+ эритроцитов на ионы К+ плазмы крови в соотношении 3:2. Кроме указанного трансмембранного обмена Na+/K+, Na+ помпа осуществляет еще, по крайней мере, четыре транспортных процесса: Na+→ Na+ обмен; K+→K+обмен; одновалентный вход ионов Na+, сопряженный с выходом К+.
Молекулярной основой Na+ помпы является фермент Na+, K+ –АТФ-аза – интегральный белок, прочно связанный с мембранными липидами, состоящий из 2х полипептидных субъединиц с ММ 80-100кД.
Транспортная система имеет 3 центра, связывающих ионов Na+, локализованных на цитоплазматической стороне мембраны. С наружной стороны мембраны на транспортной системе имеется 2 центра связывания ионов К+. Важная роль в поддержании высокой активности фермента отводится мембранным фосфолипидам.
Функционирование Са2+-помпы обеспечивается нуклеотидами, а также макроэргическими соединениями, преимущественно АТФ, ЦТФ, ГТФ, в меньшей степени ГТФ и ЦТФ.
Как в случае Nа+-помпы, функционирование Са2+помпы в эритроцитах связано с проявлениями активности Са2+, Mg2+ –АТФ-азы. В мембране одного эритроцита обнаруживается около 700 молекул Са2+, Mg2+ –АТФ-азы.
Наряду с барьерной и транспортной функциями, мембрана эритроцитов выполняет рецепторную функцию.
Экспериментально доказано наличие на мембране эритроцитов рецепторов к инсулину, эндотелину, церулоплазмину, а2-макроглобулину, α- и β-адренорецепторов. На поверхности эритроцитов находятся рецепторы к фибриногену, обладающие достаточно высокой специфичностью. Эритроциты также несут на мембране рецепторы к гистамину, ТхА2, простациклину.
В мембране эритроцитов обнаруживаются рецепторы для катехоламинов, снижающих подвижность жирных кислот липидов мембран эритроцитов, а также осмотическую устойчивость эритроцитов.
Установлена перестройка структуры мембраны эритроцитов под влиянием низких концентраций инсулина, гормона роста человека, простагландинов группы Е и Е2.
В мембранах эритроцитов высока и ц – АМФ активность. При увеличении концентраций в эритроцитах ц–АМФ ( до 10–6 М) усиливаются процессы фосфорилирования белков, что приводит в свою очередь к изменению степени фосфорилированности и проницаемости мембран эритроцитов для ионов Са2+.
Эритроцитарная мембрана содержит изоантигены различных систем иммунологических реакций, определяющих групповую принадлежность крови человека по этим системам.
4.4. Антигенная структура эритроцитарной мембраны
Эритроцитарная мембрана содержит различные антигены видовой, групповой и индивидуальной специфичности. Различают два вида изоантигенов эритроцитов, определяющих групповую специфичность крови человек – А и В агглютиногены. Соответственно в плазме или сыворотке крови обнаруживаются две разновидности изоантител – агглютинины α и β. В крови человека не содержатся одноименных агглютиногенов и агглютининов. Их встреча и взаимодействие может возникать при переливании несовместимых групп крови, приводить к развитию агглютинации и гемолиза эритроцитов.
Как известно, I (0) группа крови характеризуется отсутствием в эритроцитах агглютиногенов А и В при наличии в плазме или сыворотке крови агглютининов α и β, встречается у 40-50 % людей стран центральной Европы.
II (А) группа крови характеризуется наличием в мембране эритроцитов агглютиногена А, в то время как в плазме крови содержатся агглютинины β. Указанная группа крови распространена у 30–40 % людей.
III (В) группа крови характеризуется наличием агглютиногена В в мембране эритроцитов, а в плазме или сыворотке крови – наличием агглютининов типа α. Эта группа крови имеет место примерно у 10 % населения.
IV (АВ) группа крови характеризуется наличием в мембране эритроцитов фиксированных А и В агглютиногенов, при этом в плазме или сыворотке крови отсутствуют естественные агглютинины α и β. Данная группа крови встречается у 6 % населения.
Генетический контроль антигенной системы А,В,О мембран эритроцитов представлен генами О, Н, А, В, локализованными в длинном плече 9-й пары хромосом.
Агглютинины α и β относятся к классу Ig M, являются естественными антителами, образуются у ребенка на первом году жизни, достигая максимума к 8 – 10 годам.
Второе место среди антигенных свойств мембран эритроцитов по клинической значимости занимает система Rh – Hr. Впервые Резус-фактор был открыт в 1940 году К. Ландштейнером и А. Винером, содержится в эритроцитах у 85 % людей белой расы. У 15 % людей эти эритроцитарные антигены отсутствуют. В настоящее время установлена липопротеидная природа антигенов данной системы, их насчитывается около 20, они образуют различные комбинации в мембране эритроцитов. Наиболее распространенными резусантигенами являются 6 разновидностей: Rh0 (D), rh’ (C), rh’’ (E), Hr0 (d), hr’ (c), hr’’ (e). Наиболее сильным антигеном этой группы является Rh0 (D).
Антитела системы Rh и Hr – антирезусагглютинины являются приобретенными, иммунными, отсутствуют в крови Rh (-) людей с момента рождения, синтезируются при первом переливании Rh (+) крови Rh (-) реципиенту, а также при первой беременности Rh (-) женщины Rh(+) плодом. При первой беременности эти антитела синтезируются медленно в течение нескольких месяцев в небольшом титре, не вызывая серьезных осложнений у матери и плода. При повторном контакте резус-отрицательного человека с резус-положительными эритроцитами возможен резус-конфликт. Антитела системы Rh – Hr относятся к классу Ig G, поэтому они легко проникают через плацентарный барьер, вызывают реакции агглютинации и гемолиза эритроцитов плода, что сопровождается развитием гемолитической желтухи новорожденных. В случае повторного переливания несовместимой по Rh–антигенам крови донора и реципиента может наблюдаться гемотрансфузионный шок.